Multicellulær kompleksitet og kroppsplan
Bilde 1. Cellefabrikken
 Biokjemi handler i utgangspunktet om instruksjonell kompleks informasjon og spesifikasjoner som styrer fremstillingen og kontrollen av komplekse molekylær-fysiologiske strukturer, som dikterer funksjon.
Biokjemi handler i utgangspunktet om instruksjonell kompleks informasjon og spesifikasjoner som styrer fremstillingen og kontrollen av komplekse molekylær-fysiologiske strukturer, som dikterer funksjon.
Molekylære maskiner arbeider med utsøkt presisjon, nøyaktighet og spesifisitet, sammensatt av en intrikat ordnet samling av underenheter, hver med en klart definert rolle - som mekanisk låser hverandre i en bestemt tidssekvens til en unik konfigurasjon som gjør at de kan utføre sin funksjon på en effektiv og forutsigbar måte. Proteinsammenstillinger inneholder svært koordinerte bevegelige deler.
Innenfor hver proteinsammenstilling er ikke bare intermolekylære kollisjoner begrenset til et lite sett med muligheter, men reaksjon C avhenger av reaksjon B, som igjen avhenger av reaksjon A - akkurat som den ville gjort i en maskin, i vår felles erfaring. Proteiner er svært dynamiske enheter som viser svært høye grader av fleksibilitet, alt fra enkle sidekjede-rotasjoner til fullstendige omorganiseringer av deres sekundære struktur.
Å forklare organismeform avhenger av å forklare hvordan organer, vev og celler dannes og får form. På det laveste nivået i hierarkiet avhenger dannelse av celler i en flercellet organisme av spesifikasjonen av:
1. Morfogenese av eukaryote celler, struktur og form -lenke; (Se lenger ned på siden!)
2. Bestemmelse og differensiering av celleskjebne (fenotype, eller hvilken celletype hver enkelt vil bli) -lenke.
3. Cellevekst og størrelse -lenke
4. Utvikling og celledelings-telling: Celler må programmeres til å slutte å replikere seg selv etter riktig antall celledelinger -lenke.
Bilde 2. Celle-utvikling. Takk til Otangelo Grasso
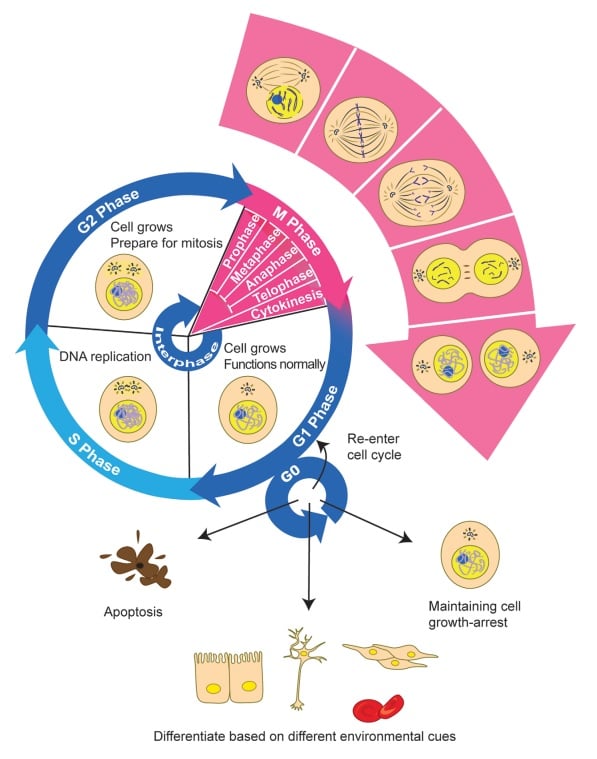 5. Mekanismer for form- dannelse -lenke.
5. Mekanismer for form- dannelse -lenke.
6. Hox-gener -lenke.
7. Posisjon og plass i kroppen. Dette er avgjørende. Lemmer som ben, finner, øyne osv. må alle plasseres på rett sted -lenke.
8. Hvilken kommunikasjon det krever å kommunisere med andre celler, og oppsettet av kommunikasjonskanalene -lenke.
9. Sensoriske og stimuli funksjoner av celler -lenke.
10. Hvilke spesifikke nye regulatoriske funksjoner celler må tilegne seg -lenke; (Lenger ned på siden)
11. Når vil utviklingsprogrammet til organismen uttrykke genene for å fremme de nye cellene under utvikling? -lenke
12. Endring av regulering i sammensetningen av cellemembranen og/eller utskilte produkter.
13. Spesifikasjon av celle-celle-adhesjonsproteinene og hvilke som skal brukes i hver celle for å feste seg til nabocellene (det er 4 klasser)
14. Apoptose: programmering av tidsperioden cellen holdes i live i kroppen, og når er det på tide å selvdestruere og erstattes av nyproduserte celler av samme type
15. Sett opp hver celles spesifikke ernæringsbehov 16. Celleform-endringer
Bilde 3. Cellens kjemi. Takk til Otangelo Grasso
 17. Celleproliferasjon er prosessen som resulterer i en økning i antall celler, og er definert av balansen mellom celledelinger og celletap gjennom celledød eller differensiering.
17. Celleproliferasjon er prosessen som resulterer i en økning i antall celler, og er definert av balansen mellom celledelinger og celletap gjennom celledød eller differensiering.
18. Forskjeller i regulatorisk DNA
Hver celle i et embryo mottar molekylære signaler fra naboceller i form av proteiner, RNA og til og med overflateinteraksjoner. Nesten alle dyr gjennomgår en lignende sekvens av hendelser under veldig tidlig utvikling, en konservert prosess kjent som embryogenese. Under embryogenese eksisterer celler i tre kimlag, og gjennomgår gastrulering. Et grunnleggende sett av de samme proteinene og mRNAene er involvert i embryogenese.
Hvordan oppsto komplekse flercellede organismer, og hvordan blir genetisk og epigenetisk informasjon oversatt til en romlig organisert kroppsplan? For å tillate bygging av komplekse organer med intrikate mønstre av cellulær spesialisering, tas slike beslutninger på et overcellulært nivå på en celle-ikke-autonom måte. Organismer krever ikke bare utviklingsmekanismer, men også strategier for å arrangere og differensiere cellene sine for overlevelse og reproduksjon.
En kropp er mer enn en samling av tilfeldig distribuerte celletyper. Utvikling involverer ikke bare differensiering av celler, men også deres organisering i flercellede arrangementer som vev og organer. Når vi observerer den detaljerte anatomien til et vev som den nevrale netthinnen i øyet, ser vi et intrikat og presist arrangement av mange typer celler. Hvordan kan materie organisere seg slik at den skaper en kompleks struktur som et lem eller et øye?
Bilde 4. Øyets oppbygning og funksjon
Det er fem hovedspørsmål for embryologer som studerer morfogenese:
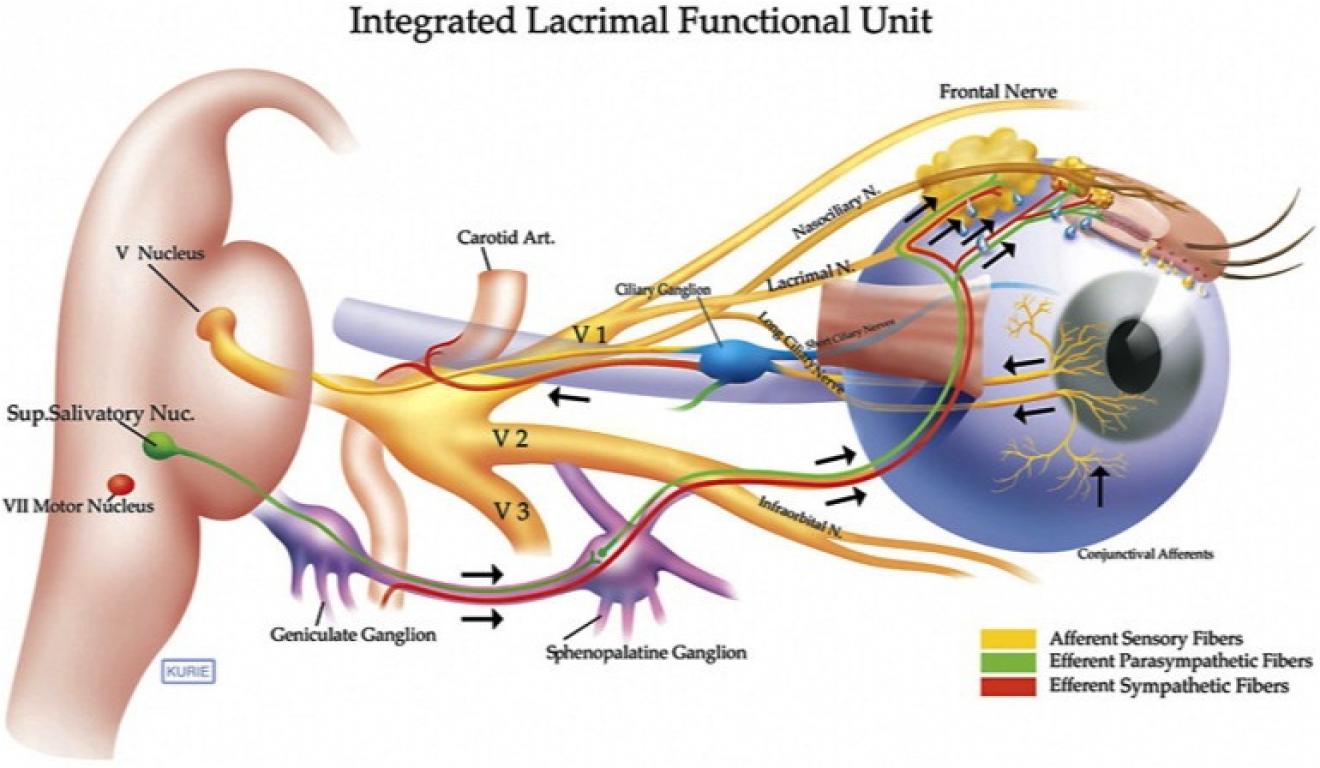 1. Hvordan dannes vev fra populasjoner av celler? For eksempel, hvordan fester nevrale netthinnen-celler seg til andre nevrale netthinnenceller uten å bli integrert i den pigmenterte netthinnen eller iris-cellene ved siden av dem? Hvordan er de ulike celletypene i netthinnen (de tre distinkte lagene av fotoreseptorer, bipolare nevroner og ganglionceller) ordnet slik at netthinnen er funksjonell?
1. Hvordan dannes vev fra populasjoner av celler? For eksempel, hvordan fester nevrale netthinnen-celler seg til andre nevrale netthinnenceller uten å bli integrert i den pigmenterte netthinnen eller iris-cellene ved siden av dem? Hvordan er de ulike celletypene i netthinnen (de tre distinkte lagene av fotoreseptorer, bipolare nevroner og ganglionceller) ordnet slik at netthinnen er funksjonell?
2. Hvordan er organer bygget opp av vev? Netthinnen i øyet dannes i nøyaktig avstand bak hornhinnen og linsen. Netthinnen ville være ubrukelig hvis den utviklet seg bak et bein eller midt i nyren. Dessuten må nevroner fra netthinnen komme inn i hjernen for å innervere områdene i hjernebarken som analyserer visuell informasjon. Alle disse koblingene må bestilles nøyaktig.
3. Hvordan dannes organer på bestemte steder, og hvordan når migrerende celler sine destinasjoner? Øyne utvikler seg bare i hodet og ingen andre steder. Hva hindrer et øye i å dannes i et annet område av kroppen? Noen celler for eksempel, forløperne til våre pigmentceller, kjønnsceller og blodceller må reise lange avstander for å nå sine endelige destinasjoner. Hvordan blir celler instruert til å reise langs bestemte ruter i våre embryonale kropper, og hvordan får de beskjed om å stoppe når de har nådd sine riktige destinasjoner?
Bilde 5. Fosutvikling menneske
 4. Hvordan vokser organer og deres celler, og hvordan er deres vekst koordinert gjennom utviklingen? Cellene i alt vevet i øyet må vokse på en koordinert måte hvis man skal se. Noen celler, inkludert de fleste nevroner, deler seg ikke etter fødselen. I motsetning til dette, avgir tarmen stadig celler, og nye tarmceller regenereres hver dag. Den mitotiske hastigheten til dette vevet må reguleres nøye. Hvis tarmen genererte flere celler enn den støter bort, kan det produsere tumorøse utvekster. Hvis den produserte færre celler enn den kutter ut, ville den snart bli ufunksjonell. Hva styrer raten av mitose i tarmen?
4. Hvordan vokser organer og deres celler, og hvordan er deres vekst koordinert gjennom utviklingen? Cellene i alt vevet i øyet må vokse på en koordinert måte hvis man skal se. Noen celler, inkludert de fleste nevroner, deler seg ikke etter fødselen. I motsetning til dette, avgir tarmen stadig celler, og nye tarmceller regenereres hver dag. Den mitotiske hastigheten til dette vevet må reguleres nøye. Hvis tarmen genererte flere celler enn den støter bort, kan det produsere tumorøse utvekster. Hvis den produserte færre celler enn den kutter ut, ville den snart bli ufunksjonell. Hva styrer raten av mitose i tarmen?
5. Hvordan oppnår organer polaritet? Hvis man skulle se på et tverrsnitt av fingrene, ville man se en viss organisert samling av vevsbein, brusk, muskler, fett, dermis, epidermis, blod og nevroner. Ser man på et tverrsnitt av underarmen, vil man finne den samme samlingen av vev. Men de er ordnet veldig forskjellig i forskjellige deler av armen. Hvordan har det seg at de samme celletypene kan ordnes på forskjellige måter i forskjellige deler av samme struktur? Alle disse spørsmålene gjelder aspekter ved celleadferd. Det er to hovedtyper av cellearrangementer i embryoet: epitelceller, som er tett koblet til hverandre i ark eller rør, og mesenkymale celler, som ikke er koblet til hverandre og som fungerer som uavhengige enheter. Morfogenese frembringes gjennom et begrenset repertoar av variasjoner i cellulære prosesser innenfor disse to typene arrangementer:
Bilde 6. Én epigenetisk mekanisme
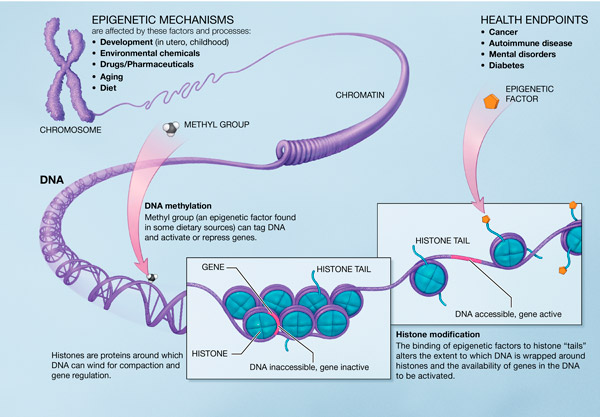 Å svare på spørsmålene om hvordan celler, vev og organismer dannes, går foran spørsmålet om hvordan de til slutt kan variere, utvikle seg, endre og forvandle seg fra en art til en annen gjennom en makroevolusjonær overgangssone for primære arter, der nye organismetrekk oppstår, som vinger, øyne, ører, ben, armer og så videre. Faktum er at vitenskapen fortsatt er LANGT fra å kunne svare på det spørsmålet på en uttømmende måte. Minst 43 epigenetiske koder og språk er vitenskapelig kjent. De bidrar alle til organismeutvikling på en avgjørende måte. Et Google-søk vil vise at det ikke er noen vitenskapelige artikler som viser løsning for bare én av disse kodene. Vitenskapen har INGEN ANNING om hvordan og hvor de fleste av dem er lagret, heller ikke betydningen av kodifiseringen, oppgaven og reglene, språket eller sifrering. Vitenskapen vet at de eksisterer, gjennom eksperimentelle tester, men dypere detaljer er ikke kjent.
Å svare på spørsmålene om hvordan celler, vev og organismer dannes, går foran spørsmålet om hvordan de til slutt kan variere, utvikle seg, endre og forvandle seg fra en art til en annen gjennom en makroevolusjonær overgangssone for primære arter, der nye organismetrekk oppstår, som vinger, øyne, ører, ben, armer og så videre. Faktum er at vitenskapen fortsatt er LANGT fra å kunne svare på det spørsmålet på en uttømmende måte. Minst 43 epigenetiske koder og språk er vitenskapelig kjent. De bidrar alle til organismeutvikling på en avgjørende måte. Et Google-søk vil vise at det ikke er noen vitenskapelige artikler som viser løsning for bare én av disse kodene. Vitenskapen har INGEN ANNING om hvordan og hvor de fleste av dem er lagret, heller ikke betydningen av kodifiseringen, oppgaven og reglene, språket eller sifrering. Vitenskapen vet at de eksisterer, gjennom eksperimentelle tester, men dypere detaljer er ikke kjent.
Det naive synet fra uinformerte mennesker er at evolusjon er et faktum og at mikroevolusjon fører til makroevolusjon, og at gener er nok til å forklare fenotype og fysiologisk form. Det er, som ovenfor vist, usant. Variasjon innen arter på andregrads-nivå, som til og med fører til artsdannelse der innavl ikke lenger er mulig, er ikke bevis på evolusjon, men forhånds-programmert tilpasning, som er livsviktig. Da livet startet, måtte tilpasningen være fullt operativ, ellers ville organismen snart dø. Det forklarer hvorfor svært varierte organismeformer innen arter kan oppstå i løpet av få generasjoner. Hvorfor kunstig avl kan produsere en grand-danois hund og en chihuahua.
Bilde 7. Eks. på én protein-sammensetning
Grasso skisserer mekanismene involvert her:
 Påstand: I biokjemi er det bare kjemiske reaksjoner.
Påstand: I biokjemi er det bare kjemiske reaksjoner.
Svar: Bruce Alberts: "The Cell as a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists," Cell, 92 (6. februar 1998): 291-294)
"Vi har alltid undervurdert celler. Det gjør vi utvilsomt fortsatt i dag. Men vi er i hvert fall ikke lenger så naive som vi var da jeg var hovedfags-student på 1960-tallet. Da så de fleste av oss på at celler inneholdt et gigantisk sett med andreordens reaksjoner: molekylene A og B ble antatt å diffundere fritt, tilfeldig kollidere med hverandre for å produsere molekyl AB - og likeledes for de mange andre molekylene som interagerer med hverandre inne i en celle.,,,
Men i stedet for en celle dominert av tilfeldig kolliderende individuelle proteinmolekyler, vet vi nå at nesten alle store prosesser i en celle utføres av sammenstillinger av 10 eller flere proteinmolekyler. Og når den utfører sine biologiske funksjoner, samhandler hver av disse protein-sammenstillingene med flere andre store komplekser av proteiner. Hele cellen kan faktisk sees på som en fabrikk som inneholder et forseggjort nettverk av sammenlåsende samlebånd, som hver er sammensatt av et sett med store proteinmaskiner."
Hvorfor Darwins evolusjonsteori ikke forklarer biologisk mangfold -lenke.
Vi vet som et faktum at felles avstamning er en mislykket hypotese:
Felles avstamning, livets tre, en mislykket hypotese -lenke.
Det forklarer imidlertid ikke på LANGT nær økningen av flercellet kompleksitet og biologisk variasjon av arter. For det må det molekylære landskapet være mer komplekst og involvere mange forskjellige mekanismer på intra- og ekstra-cellulært nivå som forklart ovenfor, og her:
Hvor kommer komplekse organismer fra? -lenke.
Oversettrelse og bilder ved Asbjørn E. Lund